Taste preference thresholds
The recorded taste preference threshold values for each of the five carbohydrates in white-faced sakis are the first assessment of taste responsiveness in a frugivorous seed predator and fall generally within the lower range of those reported in other primates. Contrary to the initial hypotheses that the sakis would show a low taste sensitivity towards sweet-tasting carbohydrates due to their sclerocarpic behavior, the results showed similar taste responsiveness to that of other ripe fruit specialists. It was unexpected to find that the white-faced sakis showed overlapping taste preference thresholds to those reported in other frugivorous primates: first, due to their unique dietary specializations and second, because sakis avoid competition with ripe fruit predators by consuming low-carbohydrate food items such as fruits in earlier ripening stages.
In general, it is conceivable that the resemblance observed in the taste preference thresholds for the tested carbohydrates in white-faced sakis and other frugivorous primates could be linked to the general adaptation to a frugivorous diet, despite the possibility of slight variations in palatability reflecting differences in dietary adaptations.
Table 1. Taste preference threshold ranges for food-associated carbohydrates and steviol glycosides in primates (mM)

Sucrose & Fructose
The taste preference threshold values for sucrose and fructose scored by the white-faced sakis fall generally within the threshold ranges established in primates thus far studied (Table 1). The sakis’ threshold values situate in the lower range of values among not only platyrrhines but primates, just above Ateles geoffroyi (3 mM for sucrose; 15 mM for fructose), and abreast to those of Saimiri sciureus (10 mM sucrose; 40mM fructose). The sakis also showed similar taste preference thresholds for sucrose and fructose as other non-human primate species in which evidence for seed predation has been reported.
For fructose, the threshold values displayed by the sakis are placed among the lowest values scored by any primates studied so far. Specifically, the threshold value of 10 mM scored by two individuals is qualitatively lower than that reported in any other platyrrhine, strepsirrhine, and hominoid studied to date, outscoring the second lowest threshold of 15 mM for this carbohydrate reported for the Bornean orangutan and Geoffroy´s spider monkey.
Glucose, Maltose & Lactose
The threshold values for glucose found in the white-faced sakis (20-30 mM) range among the lowest ones reported so far in primates (20-330 mM), indicating a high taste sensitivity. These values compare to those found in Cercopithecidae, ranging between 20-25 mM, far from those scored by the prosimian Varecia variegata (75 mM), or Pan troglodytes and humans (both with 80 mM). Free glucose concentration in ripe fruits ranges between 94.4-1378.1 mM/100 g of carbohydrates, and thus, is clearly perceptible for the sakis. In addition, the taste preference thresholds for maltose scored by white-faced sakis (20-30 mM) fall into the threshold range scored by the other studied non-human primates, varying between 10-90 mM.
White-faced sakis exhibit a lower taste preference threshold for lactose compared to other primate species. The lactose content of the milk of white-faced sakis is yet unstudied, and lactose content of Pithecia milk have yielded no corresponding data. However, it is reasonable to assume that the lactose content in sakis milk is not fundamentally different from that of other New World primates. The lactose content in primate milk typically ranges between 175-263 mM, which is substantially higher than the threshold values determined in the present study (30-40 mM), and thus, clearly perceptible for the sakis.
The preference for sweet-tasting substances among mammals may be linked to their exposure to the sweet taste of lactose in milk, which is the primary source of nutrition for mammals during their early life. Following this hypothesis, it would be plausible to predict a correlation between the lactose content in milk and an animal´s sweet-taste sensitivity, as it has been shown in strepsirrhines. Taking sucrose as a representative of sweet-taste sensitivity, the results of the current study demonstrate a strong positive correlation between taste preference thresholds for sucrose and lactose in both platyrrhines and in primates (Spearman, rs = 0.975, p < 0.01; Spearman, rs = 0.868, p < 0.01), thereby supporting the previous hypothesis.
Steviol glycosides
Taste preference thresholds for rebaudioside A and stevioside have been established only in a few primate species, with the sakis being the first tested frugivorous seed predator. As in the case with the taste preference threshold for food-associated carbohydrates, the preference threshold values for rebaudioside A and stevioside in white-faced sakis (0.04 and 0.2-0.5 mM, respectively) fall within the range of those reported in other primates (Table 1). These values are lower than the reported concentrations of rebaudioside A (21.5 mM) and stevioside (11.2 mM) in fresh leaves of Stevia rebaudiana making both steviol glycosides clearly perceptible for the sakis when feeding on this neotropical plant.
The comparable taste preferences for these sweeteners between the tested primate species to date might imply that frugivorous and omnivorous primates possess similar sweet-taste receptors that exhibit a comparable response to high-potency sweeteners.
Relative taste preferences
The pattern of relative preference for food-associated carbohydrates of white-faced sakis is consistent with those found in other platyrrhines, prosimians, and hominoids (including humans). Although white-faced sakis are considered seed predators whereas these other species act as seed dispersers, all of them rely heavily on fruits, preferring sucrose over all other carbohydrates and perceiving fructose as the second-most attractive carbohydrate.
The differences in the attractiveness of sweet-tasting carbohydrates are thought to reflect the differences in their stimulating efficiency at equimolar concentrations. As a result, the relative taste preferences of white-faced sakis mirror the experienced relative sweetness of these substances. Interestingly, the order of the taste preference thresholds displayed by the sakis for the five carbohydrates tested here is similar to the order of relative preferences exhibited when presented with suprathreshold concentrations of these sweet-tasting substances. The lowest preference thresholds scored by the sakis correspond to sucrose (10 mM) and fructose (10-40 mM) which are the most and second-most preferred carbohydrates, respectively. In contrast, lactose had the highest preference thresholds (30-40 mM) and was placed as the least preferred carbohydrate. However, the pattern of relative taste preferences found in the white-faced sakis were not consistent with the pattern of sweetness potency of the tested food-associated carbohydrates: fructose > sucrose > glucose > maltose > lactose.
The pattern of relative sweetness was identical both at 100 mM and 200 mM (Sucrose > fructose > glucose = maltose = lactose), whereas at 300 mM the white-faced sakis additionally showed a clear preference for glucose over maltose as a group (sucrose > fructose > glucose > maltose = lactose). Since the sakis´ taste preference thresholds were higher for glucose than for sucrose, it is reasonable to assume that higher concentrations of glucose were needed to obtain a similar stimulating efficiency, as described in two neotropical frugivorous primates, Saimiri sciureus and Ateles geoffroyi or in Pan troglodytes. These findings are consistent with the finding that sugars with presumably similar sweetness can elicit distinct preferential responses due to the differences in sweetening potency between sugars at different concentration level.
Taste sensitivity and body mass
Larger primates are more likely to include a greater variety of foods containing lower sugar concentrations into their diet to cope with higher basal metabolic rates and thus high energy demands. In contrast, despite the lower basal metabolic rates and thus lower energy requirements, smaller frugivorous primates experience costlier energy expenditures when foraging. Therefore, smaller species may benefit from a higher sensitivity to sweet-tasting carbohydrates as a way of efficiently detecting energy-rich food resources.
Body mass and taste preference thresholds sucrose in primates were significantly correlated when including the results from the present study to the available data primates tested to date (Pearson, r = -0.67, p < 0.001) (Fig. 4). In contrast, the correlation between body mass and taste preference thresholds for fructose in primates was not statistically significant (Pearson, r = -0.02, p > 0.1) (Fig. 4).
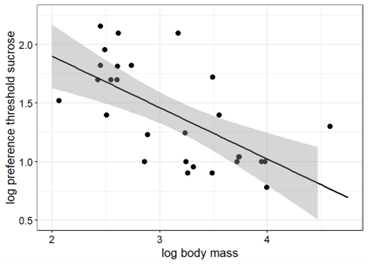
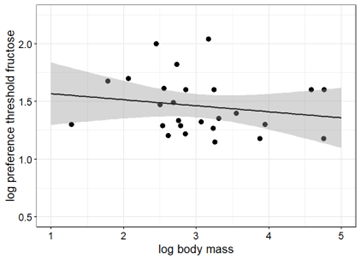
Figure 4. Taste preference thresholds for sucrose (left) and fructose (right) in 32 and 27 primate species as a function of body mass (all logarithms to base 10). The regression with the best goodness-of-fit inferred from the linear regression analysis is represented by the solid line (sucrose: R2 = 0.45, y = 2.92 – 0.48x; fructose: R2 = 0.001; y = 1.5 – 0.01x).
When considering platyrrhines alone, the correlation between body mass and taste preference thresholds for sucrose was statistically significant (Pearson, r = -0.678, p < 0.05) whereas it was not for fructose (Pearson, r = -0.438, p > 0.1, respectively). Figure 5 illustrates a negative correlation between taste preference thresholds for sucrose (Fig. 5) and fructose (Fig. 5) and body mass in platyrrhine primates, including the data of white-faced sakis obtained in the current study.
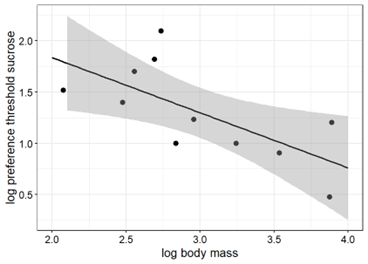
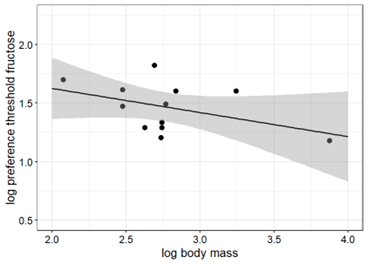
Figure 5. Taste preference thresholds for sucrose (left) and fructose (right) in 11 and 12 platyrrhine primate species as a function of body mass (all logarithms to base 10). The regression with the best goodness-of-fit inferred from the linear regression analysis is represented by the solid line (sucrose: R2 = 0.46, y = 2.91 – 0.54x; fructose: R2 = 0.19; y = 2.04 – 0.21x).
The findings of the present study support the proposed correlation between body mass and taste preference thresholds for sucrose in primates (Fig. 4A, 5A). In contrast, the correlation between body mass and taste preference thresholds for fructose is not statistically significant (Fig. 4B, 5B), perhaps due to the limited amount of data.
The average weight of adult white-faced sakis ranges between 1.5-4.0 kg and therefore, the relatively low taste preference thresholds for sucrose and fructose of the sakis fit to the notion of the presumed correlation between body mass and taste preference thresholds for these carbohydrates when compared to other larger primates. However, other smaller platyrrhines show higher taste preference threshold scores for these carbohydrates than those reported in white-faced sakis, contradicting the suggested correlation. This could be explained by the lack of correlation between body size and the degree of seed predation. As a result, sakis might exhibit exceptional taste preference thresholds for their body size.
Taste responsiveness and seed predation
The results of the present study support the suggestion that pitheciids, and particularly white-faced sakis might represent Liem’s paradox in primates. In line with this, the seasonal diversification of the diet and changes in the degree of ripening and hardness of consumed seeds and fruits mirror the extension of the range of suitable food items exploited by the white-faced sakis even if lipid-rich seeds are considered as their principal resource of metabolic energy consumed year-round. This expansion of the sakis´ dietary niche mirrors the behavioural adaptation to minimize the effects of the seasonal resource availability, consistent with the optimal foraging theory.
The seasonal changes in the feeding habits of white-faced sakis appear to be closely linked to the ripening stages of the consumed resources. Sakis predate fruits in different states of ripening, and thus, of different nutritional compositions. Sakis might track the changes in the content of free simple sugars to assess lipid and protein contents in consumed fruits, highlighting the relevance of their high taste sensitivity for food-associated carbohydrates for selecting the most beneficial resources and rejecting the least unpalatable ones. In addition, the observed high taste sensitivity for detecting carbohydrates might have resulted in an adaptation to avoid not only seasonal ripe fruit scarcity but competition with larger primate species by being able to consume fruits at earlier ripening stages.


Therefore, the obtained taste preference thresholds for food-associated carbohydrates appear to be in discordance with the sakis´ seed predation and the high lipid consumption. The positive response to the sweet taste of sugars reported in the present study could manifest a specific adaptation to efficiently access and exploit energy-rich foods such as ripe fruits, consistent with the optimal foraging theory. In addition, the relative preference pattern for sweet-tasting carbohydrates obtained in white-faced sakisis similar to that reported in other frugivorous species. However, such taste responsiveness for food-associated carbohydrates might reflect the ability of white-faced sakis to detect low sugar concentrations in seeds and unripe fruits, and thus presents an advantage concerning the highly valuable resources exploited by other frugivorous primates at earlier ripening stages and overcome the seasonal fluctuation in resource availability.