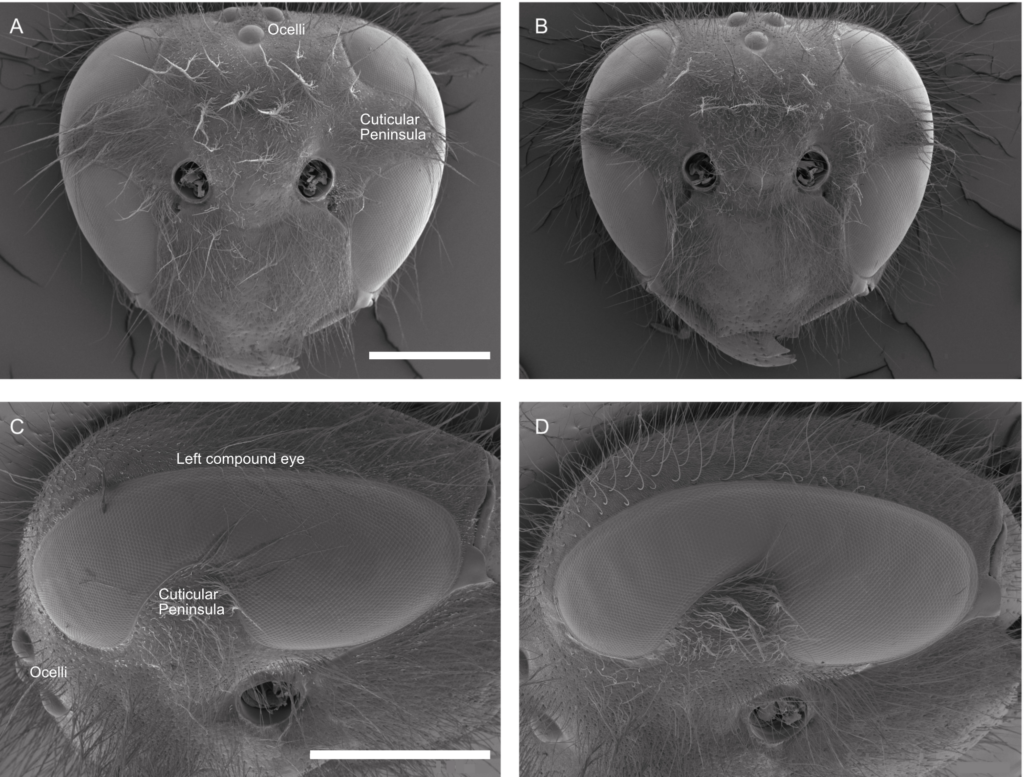
In the compound eyes of insects, the density of the ommatidia is represented by the interommatidial angle (Δφ), which defines the anatomical resolution of the eye. If an individual has a smaller Δφ, it will potentially have a greater spatial resolution. A possible complicating factor is that these and many other wasps possess an equatorial cuticular “peninsula” that partially divides their compound eyes into dorsal and ventral halves. Even though this peninsula potentially could have created a blind spot in the frontal visual field, I discovered that this was not the case in the studied two wasps (Figure 1). Interestingly, however, how interommatidial angles varied within the eye changed abruptly at this peninsula, with the dorsal half of the eye generally having higher Δφ and lower spatial resolving power than the ventral half of the eye.
In general, I found that the average interommatidial angle in wasps decreases smoothly from the dorsal part of the eye to the equator and increases smoothly again towards the ventral part of the eye (latitude from 0° to -80°). Although both species exhibited small Δφ values, V. germanica showed the smallest average Δφ of 1.0°, at around -10° latitude and between 10 and 20° in longitude. Although V. vulgaris has the larger minimum values of Δφ, they are found in an eye region similar to that in V. germanica. These small values of Δφ (Fig. 7), together with narrower photoreceptor receptive fields at the same location, indicate the presence of an “acute zone” that allows higher spatial resolution in the frontal-ventral part of the eye. The acute zones of these wasps afford a visual performance rivalling the best seen in other day-flying hymenopterans (e.g. the carpenter bees X. leucothorax and X. tenuiscapa)
If I compare the photoreceptor receptive fields of V. germanica and V. vulgaris with some other insects, it is evident that these wasps have reasonably small values of acceptance angle Δρ, and thus high spatial resolution. Bees have a frontal acute zone within which photoreceptors have a frontal average acceptance angle of 1.6°, while other species such as the drone fly Eristalis tenax (Δρ = 0.9°) and the butterfly Melanitis leda (Δρ = 1.5°) also have similar acceptance angles and consequently similar spatial resolution.
Insects with Δρ less than about 1° are often predators, including dragonflies, mantids, killer flies and some wasps, among others. These insects typically have a forward pointing acute zone for tracking prey. This offers one explanation for the small Δφ measured for the wasps used in the present study. V. germanica and V. vulgaris are predators and scavengers that seek different types of protein sources, including flies, lepidopteran larvae, ants, grasshoppers and arachnids, among others. Both species hunt from above, and their frontal-ventral acute zones would thus be beneficial for this purpose. Floral nectar is also an important part of the diet of these wasps and their acute zones are well positioned to view flower structure as the wasps land. Additionally, V. germanica and V. vulgaris are ground-nesting wasps and the nest entrance is well positioned within the acute zone during landing.
When I calculated the optical sensitivity S of the eyes of the two wasps, I found that both have values typical of diurnal insects with apposition compound eyes: 0.20 µm2sr for V. germanica and 0.14 µm2sr for V. vulgaris. These values are comparable to those calculated for other diurnal hymenopterans having apposition eyes, for example, the carpenter bees Xylocopa tenuiscapa and Xylocopa leucothorax (S = 0.3 and 0.1 µm2sr, respectively), the honeybee Apis mellifera (S = 0.1 µm2sr) and the wasp Polistes occidentalis (S = 0.1 µm2sr). In contrast, the optical sensitivities of nocturnal bees and wasps tend to be around 10-30 times higher, as might be expected of eyes adapted for a life in dim light.
In conclusion, V. germanica and V. vulgaris have compound eyes that are typical of visually active diurnal insects. They each possess an acute zone in the ventral-frontal part of the eye, with high spatial resolution subserved by small interommatidial angles and narrow photoreceptor receptive fields (i.e. low acceptance angles), optical properties well suited to the hunting and foraging lifestyles that characterise these two species of wasps.